



Three Theories of Morphogenesis (Mechanistic, Vitalist and Organismic)
By Rupert Sheldrake

The word morphogenesis means the coming-into-being of form, from the Greek morphe = form and genesis = coming-into-being. Biological morphogenesis is the coming-into-being of characteristic and specific forms in living organisms, such as the flowers of foxgloves and the ears of rabbits, and of course whole organisms themselves.
Morphogenesis is one of the great unsolved problems of biology. In this essay, I summarize why it is a problem and then look at the three main kinds of theory that have been proposed to explain it: mechanistic, vitalist and organismic.
Since the early twentieth century the mechanistic theory has been predominant within academic biology, and by the late twentieth century, the advances of molecular biology made many people optimistic that the problems would soon be solved mechanistically in terms of genes, proteins and other molecules. However, these hopes have now been dashed, or at least postponed, Growing numbers of biologists, including Michael Levin, at Tufts University in the US, argue that we need to move to a more holistic, organismic approach. I have been advocating such a shift since 1981, with the publication of my first book, A New Science of Life. This essay is based on some of the points I make in Chapter 2 of that book and anyone interested in following up in more detail or looking at the references can find them in the third edition of the book (re-named Morphic Resonance in the US).
Developmental processes can be described in many ways, but describing is not the same as understanding. The external form of the developing animal or plant can be drawn, photographed or filmed, providing a series of pictures of its changing morphology; its internal structure can be described at successive stages; changes in physical quantities such as weight, volume and rate of oxygen consumption can be measured; and changes in the chemical composition of the system as a whole and of regions within it can be analysed.

Figure: Stages in the development of the embryo of the shepherd’s purse plant, Capsella bursa-pastoris.
The progressive improvement of techniques enables such descriptions to be made in ever greater detail. With electron microscopes, the processes of cellular differentiation can be studied at higher resolution than with the light microscope, enabling many new structures to be seen. The sensitive analytical methods of molecular biology enable changes in concentrations of specific molecules, including proteins and nucleic acids, to be measured in very small samples of tissue. By means of radioactive isotopes or fluorescent antibodies, chemical structures can be “labelled” and “traced” as the system develops; and techniques for inducing genetic changes in some of the cells of embryos enable their genetically “marked” descendants to be identified and their fate to be “mapped”.
Most research in embryology and developmental biology is about descriptions that are then classified and compared. But descriptive results cannot in themselves lead to an understanding of the causes of development, although they may suggest hypotheses, which can then be investigated by means of experimental disturbances of development.
The major problems thrown up by this type of research can be summarized as follows:
The first problem is morphogenesis itself: form comes into being. New structures appear, such as eyes or flowers, which cannot be explained in terms of structures already present in the egg. There are no miniature eyes in eagles’ eggs, or miniature flowers in dandelion seeds. Biological morphogenesis involves an increase in complexity of form and organization which cannot be explained in terms of the unfolding of pre-formed but invisible structures. More form comes from less.
Many animal embryos and developing plants can regulate, in other words, produce a normal structure if part of the system is destroyed or removed at a sufficiently early stage. For example, if half of a dragonfly’s egg is separated off and killed, the remaining half goes on to produce a small but complete dragonfly embryo.
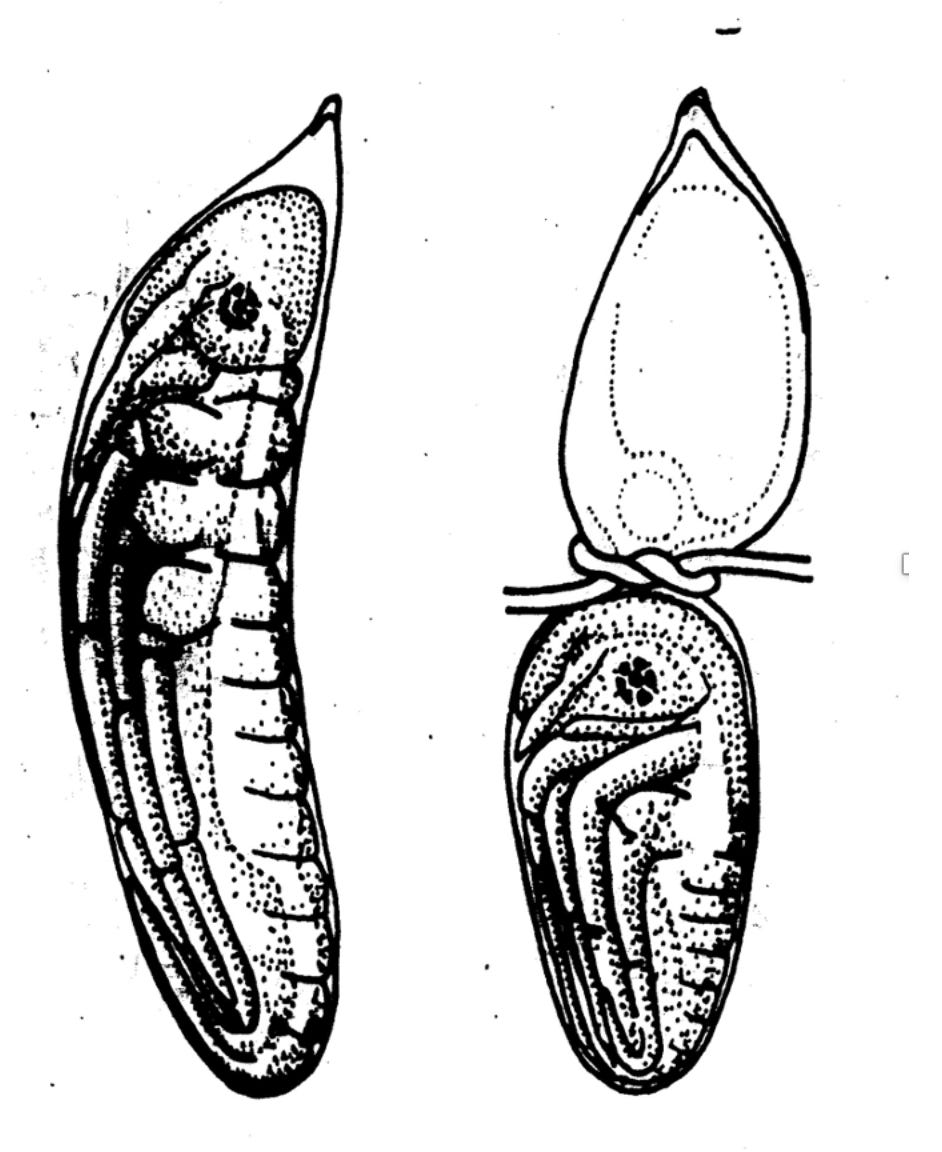
Figure: A normal dragonfly embryo (left) and a small but complete embryo that has formed from half the original egg.
Many systems can regenerate or replace missing parts. For example, a new willow tree can develop from a small cutting, and new flatworms can develop from pieces sliced out of a previous flatworm.
In vegetative and sexual reproduction, new organisms are formed from detached parts of parent organisms.
Mechanistic, vitalist and organismic theories all start from this established body of facts and results, about which there is general agreement, but they differ radically in their interpretations.
Mechanistic Theories
Mechanistic theories of morphogenesis ascribe a role of prime importance to the genetic material DNA, for four main reasons:
First, many cases of hereditary differences between animals or plants of a given species have been found to depend on genes, which can be mapped and located on chromosomes.
Secondly, the chemical basis of genes is known to be DNA, and their specificity is known to depend on the sequence of purine and pyrimidine bases in the DNA.
Thirdly, DNA acts as the chemical basis of heredity: it serves as a template for its own replication, owing to the specificity of the pairing of the bases in its two complementary strands; it also serves as the template for the sequence of amino acids in proteins. It does not play the latter role directly; one of its strands is first transcribed to give a single-stranded molecule of messenger RNA from which, the sequence of bases is read off three at a time in the process of protein synthesis. Different triplets of bases specify different amino acids, and thus the genetic code is translated into a sequence of amino acids, which are linked together to give characteristic polypeptide chains, which then fold up to give proteins.
Finally, the characteristics of a cell depend on its proteins: its metabolism and its capacities for chemical synthesis on enzymes, some of its structures on structural proteins, and the surface properties which enable it to be “recognized” by other cells on special proteins on its surface.
Within the mechanistic framework, the central problem of development and morphogenesis is seen as the control of protein synthesis. In bacteria, specific chemicals called inducers can cause specific regions of the DNA to be transcribed into messenger RNA, on which template specific proteins are then made. The classic example is the induction of the enzyme beta-galactosidase by the sugar lactose in the bacterium Escherichia coli. The “switching on” of the gene takes place through a complicated system involving a repressor protein which blocks transcription by combining with a specific region of the DNA; its binding to the DNA is greatly reduced in the presence of the chemical inducer, thus switching it on. By a comparable process, specific chemical repressors can switch off genes.
In animals and plants, a range of developmental genes are concerned with the regulation of the overall body plan and the pattern of body parts. These genes are usually called the “genetic toolkit”. The most surprising discovery of developmental biology in the 1990s was that these toolkit genes are remarkably similar, indeed almost identical, in widely different organisms. For example, the “homeobox” family of genes that affect the patterning of the body axis in fruit flies, mice and humans are very similar, and yet the body form of these organisms is obviously very different. But how do such different structures as the insect compound eye and the vertebrate lens-type eye develop when their formation is controlled by such similar, even functionally interchangeable genes?
This convergence of developmental and evolutionary biology has created a new field called evolutionary developmental biology (“evo-devo” for short). Most toolkit genes code for proteins that affect the activity of other genes involved in the developmental process and are part of “signalling pathways”. Some of them code for receptor proteins on cell surfaces that bind to specific molecules that act as signals.
In the early days of molecular biology, there seemed to be a simple, straightforward picture: one gene was transcribed into one messenger RNA molecule, which coded for one protein. But the picture has grown more complicated. Messenger RNA can be made up of pieces transcribed from different regions of the DNA and subsequently joined together in a specific way. Moreover, the synthesis of proteins is also controlled at the translational level; protein synthesis can be switched on and off even in the presence of appropriate messenger RNA.
The different proteins made by different types of cells thus depend on the way in which protein synthesis is controlled. The only way in which this can be understood mechanistically is in terms of physical and chemical influences on the cells; patterns of differentiation must therefore depend on physical and chemical patterns within the tissue, such as concentration gradients of specific chemicals called morphogens. The cells may also respond to electrical gradients, electrical or chemical oscillations, mechanical contacts between cells, or combinations of different factors. The cells must then respond to these differences in specific ways. One way of thinking about this problem is to regard these physical or chemical factors as providing “positional information” which the cells then “interpret” in accordance with their “genetic program” by switching the synthesis of particular proteins on or off.
Various aspects of the control of protein synthesis are under active investigation. Most biologists hope that the solution of this problem will provide, or at least lead towards, an explanation of morphogenesis in purely mechanistic terms.
Difficulties
In order to assess whether such a mechanistic explanation is likely, or even possible, a number of difficulties need to be considered:
The explanatory role of DNA and the so-called “genetic program” is severely restricted in its scope by the fact that both the DNA and the proteins of different species may be very similar. For example, in a detailed comparison of human and chimpanzee proteins, many are identical, and others to differ only slightly: the average human protein is more than 99 percent identical to its chimpanzee counterpart. Now that both genomes have been sequenced even more detailed comparisons are possible, but as Svante Paabo, the director of the chimpanzee genome commented, “We cannot see in this why we are so different from chimpanzees”.
Comparisons between closely related species of fruit fly in the genus Drosophila have revealed larger differences between different species of fruit fly than between humans and chimpanzees. Different species of mice are also more dissimilar from each other than humans and chimpanzees. If genes and proteins do not explain the differences between chimpanzees and us, then what does?
However, leaving aside all these problems, assume for the purpose of argument that the hereditary differences between chimpanzees and humans will indeed be explained in terms of small changes in protein structure, or small numbers of different proteins, or genetic changes which affect the control of protein synthesis or combinations of these factors.
Within the same organism, different patterns of development take place while the DNA remains the same. Consider, for example, your arms and legs: both contain identical cell types (muscle cells, connective tissue cells, etc.) with identical proteins and identical DNA. Therefore, the differences between arms and legs cannot be ascribed to DNA per se; they must be ascribed to pattern-determining factors that act differently in developing arms and legs. They also give rise to mirror-image patterns in right and left arms and legs. The precision of arrangement of the tissues - for example the joining of tendons to the right parts of the bones - shows that these patterns are established in detail and with precision. The mechanistic theory of life means that these factors must be regarded as physical or chemical, but their nature is unknown.
Even if physical or chemical factors affecting the growth of an arm, the formation of an eye or the development of an apple can be identified, this raises the question of how these factors are themselves patterned in the first place. However, assume for the purpose of argument that it might be possible to identify pre-existing physical factors that set up these patterns. Assume also that the ways in which these controlling factors are themselves controlled can be identified, in a series going right back right back to the fertilized egg from which the organism grew. Now there is the problem of regulation: if part of the system is removed, this complicated series of physico-chemical patterns must be disrupted. But somehow the remaining parts of the embryo manage to change their usual course of development and produce a more or less normal adult.
This problem is generally agreed to be extremely difficult; but let us assume once more that a mechanistic solution can be achieved.
The next problem is how this positional information brings about its effects. The simplest possibility is that the positional information is specified by a concentration gradient of a specific chemical, and cells exposed to more than a certain concentration synthesize one set of proteins, while cells exposed to concentrations below this threshold synthesize another set of proteins. Again, assume that mechanisms by which positional information can be interpreted can be identified and described in detail. Now, at the end of a chain of highly optimistic assumptions, we reach the situation in which different cells arranged in a suitable pattern make different proteins.
So far, there has been a set of one-to-one relationships: a gene is switched on by a specific stimulus; DNA is transcribed into RNA; and RNA is translated into a particular sequence of amino acids, a polypeptide chain. But now this simple causal sequence comes to an end. How do the polypeptide chains fold up into the characteristic three-dimensional structures of proteins? How do the proteins give the cells their characteristic structures? How do cells aggregate together to give tissues characteristic structures? And so on. These are the problems of morphogenesis proper: the synthesis of specific polypeptide chains provides the basis for the metabolic machinery and the structural materials on which morphogenesis depends. The polypeptide chains and the proteins into which they fold up are undoubtedly necessary for morphogenesis; but what actually determines the patterns and structures into which the proteins, cells and tissues combine? Mechanists assume that all this can be explained in terms of physical interactions. Morphogenesis takes place spontaneously, given the right proteins in the right places at the right times and in the right sequence. This is rather like saying that a house can build itself spontaneously as long as the right building materials are delivered to the building site at the right times. At this crucial stage, biology abdicates, and the problem of morphogenesis is left to spontaneous physical and chemical processes.
In the building-site analogy, the activity of the builders and a plan to which they are working are needed in addition to the starting materials. By contrast, morphogenesis must depend on spontaneous physical processes driven by spontaneous energy flows, and the systems must be self-organizing. But how?
In so far as these structures undergo spontaneous self-assembly, they resemble crystals; some of them, like viruses, can indeed be regarded as crystalline or quasi-crystalline. So, in principle they pose no more, or no less, of a problem than normal crystallization; the same sorts of physical process can be assumed to be at work.
Nevertheless, very few morphogenetic processes can be regarded as types of crystallization. They must involve a number of other physical factors; for example, the forces of surface tension influence the shapes taken up by membranes. And then some of the patterns may arise from statistically random fluctuations. Simple examples of the appearance of order through fluctuations have been studied from the point of view of irreversible or non-equilibrium thermodynamics in inorganic systems. Some patterning processes can be modelled mathematically in complex systems theory.
However, the mechanistic theory does not merely suggest physical processes play a part in morphogenesis; it asserts that morphogenesis is entirely explicable in terms of physics. But this does not necessarily mean it can be explained in terms of the known laws of physics.
In relation to biological morphogenesis a complete explanation would be achieved if a biologist supplied with the entire genome sequence of an organism details about the physical and chemical state of the fertilized egg, and of its environment, could predict in terms of the fundamental laws of physics (e.g. quantum field theory, the equations of electro-magnetism, the second law of thermodynamics, etc.)
The three-dimensional structure of all the proteins the organism would make.
The enzymic and other properties of these proteins.
The organism's entire metabolism.
The nature and consequences of all the types of positional information that would arise during its development.
The structure of its cells, tissues and organs and the form of the organism as a whole.
In the case of an animal, its instinctive behaviour.
If all these predictions could be made successfully, and if, in addition, the course of processes of regulation and regeneration could also be predicted a priori, this would indeed be a conclusive demonstration that living organisms are fully explicable in terms of the known laws of physics. But nothing of the sort can be done at present. So, there is no way of demonstrating that such an explanation is possible. It might not be. This is what is at stake in the Genome Wager.
The Genome Wager
In March 2009, I took part in a debate on “The nature of life” at the Cambridge University Science Festival with Lewis Wolpert, a leading exponent of the mechanistic theory of development. In this debate, Wolpert asserted that, given the genome of a fertilized human egg, it would not be long before we could predict from first principles, in terms of physical and chemical factors, the morphogenesis of a human being. I argued that this was not possible even in principle. We agreed to turn this disagreement into a formal wager lasting 20 years, and jointly paid for the stake, a case of fine port (Quinta do Vesuvio, 2005), which is being stored in the cellars of the British Wine Society.
As we were discussing the wording of the wager, Wolpert shifted his ground. Although he initially claimed we would soon be able to predict all the details about a person on the basis of the genome of a fertilized egg, on reflection he decided it would take more than 100 years to do this. Since our wager has a 20-year time span, he changed his prediction from a human to a mouse. Then he retreated to a chick, then a frog. Then he downgraded the prediction further, to a small nematode worm. Finally, we agreed that any multicellular animal or plant would qualify, however small or simple. The wager was published in New Scientist magazine on July 8, 2009, and reads as follows:
By 1 May 2029, given the genome of a fertilised egg of an animal or plant, we will be able to predict in at least one case all the details of the organism that develops from it, including any abnormalities
Even committed mechanists now think that Wolpert’s heirs are unlikely to win the bet (Wolpert died in 2021), if only because he ignored the important of epigenetic inheritance, which enables organisms to pass on adaptations they have acquired to their offspring without there being any changes in the genome.
Vitalism
Vitalism asserts that the phenomena of life cannot be fully understood in terms of physical laws derived only from the study of inanimate systems, but that additional causes are at work in living organisms. For example, in 1844, the chemist Liebig argued that although chemists could already produce all sorts of organic substances, and would in future produce many more, chemistry would never be in a position to create an eye or a leaf. Besides the recognized causes of heat, chemical affinity, and the formative force of cohesion and crystallization “in living bodies there is added yet a fourth cause which dominates the force of cohesion and combines the elements in new forms so that they gain new qualities - forms and qualities which do not appear except in the organism.”
Ideas of this type, although widely held, were too vague to provide an effective alternative to the mechanistic theory. Only at the beginning of the twentieth century were neo-vitalist theories were worked out in some detail. The most important vitalist theory was that of Hans Driesch, a German embryologist.
Driesch accepted that many features of living organisms could be understood in physico-chemical terms. He was well aware of the findings of physiology and biochemistry, and of the potential for future discovery. He knew that enzymes ('ferments') catalysed biochemical reactions and could do so in test tubes: “There is no objection to our regarding almost all metabolic processes inside the organism as due to the intervention of ferments or catalytic materials, and the only difference between inorganic and organic ferments is the very complicated character of the latter and the very high degree of their specification.” He knew that genes were material entities located in the chromosomes, and that they were probably chemical compounds of specific structure. He thought that many aspects of metabolic regulation and physiological adaptation could be understood along physico-chemical lines”. His opinions on these subjects have been confirmed by the subsequent advances of physiology, biochemistry, and molecular biology. Obviously, Driesch was unable to anticipate the details of these discoveries, but he regarded them as possible and compatible with vitalism.
In relation to morphogenesis, he considered that “it must be granted that a machine, as we understand the word, might very well be the motive force of organogenesis in general, if only normal, that is to say, if only undisturbed development existed, and if taking away parts of our system led to fragmental development.” But, in fact, in many embryonic systems the removal of part of the embryo is followed by regulation, whereby the remaining tissues reorganize themselves and go on to produce an adult organism of more or less normal form, as in the example of the dragonfly embryo pictured above.
The mechanistic theory attempts to account for development in terms of complex physico-chemical interactions between the parts of the embryo. Driesch argued that the fact of regulation made any such machine-like system inconceivable, because the system was able to remain a whole and produce a typical final result after damage, whereas no complex three-dimensional machine-like system could remain a whole after the arbitrary removal of parts.
This argument is open to the objection that it is, or will be at some time in the future, invalidated by advances in technology. But it does not seem to have been refuted so far. Computers, for example, have a fixed structure. They cannot regenerate; if parts of the computer are destroyed at random, they cannot be formed again by the machine itself, nor can the system go on functioning normally after the arbitrary removal of parts. The other item of modern technology that might seem relevant is the hologram, from which pieces can be removed but which can still give rise to a complete three-dimensional image. But significantly, the hologram is not a machine: it is an interference pattern in a field.
Driesch believed that the facts of regulation, regeneration and reproduction showed that there was something about living organisms that remained a whole even though parts of the organism could be removed. Something acted on them in a top-down manner but was not itself part of them. He called this holistic causal factor entelechy. He postulated that entelechy organized and controlled physico-chemical processes during morphogenesis. The genes were responsible for providing the material means of morphogenesis – the chemical substances to be ordered - but the ordering itself was brought about by entelechy. Clearly morphogenesis could be affected by genetic changes that changed the means of morphogenesis, but this would not prove that it could be explained simply in terms of genes and the chemicals to which they gave rise.
Similarly, the nervous system provided the means for the actions of an animal, but entelechy organised the activity of the brain, using it as an instrument, as a pianist plays on a piano. Again, behaviour can be affected by damage to the brain, just as the music played by the pianist is affected by damage to the piano; but this only proves that the brain is a necessary means for behaviour, as the piano is a necessary means for the pianist.
Entelechy is a Greek word whose derivation (en-telos) indicates something that bears its end or goal in itself; it “contains” the goal towards which a system under its control is directed. Thus, if a normal pathway of development is disturbed, the system can reach the same goal in a different way. Driesch considered that development and behaviour were under the control of a hierarchy of entelechies, which were all ultimately derived from, and subordinated to, the overall entelechy of the organism. As in any hierarchical system, mistakes were possible and entelechies might behave stupidly, as they do in cases of super-regeneration, when a superfluous organ is produced. But such stupidities do not disprove the existence of entelechy any more than scientific errors disprove that scientists are intelligent beings.
Driesch described entelechy as an “intensive manifoldness”, a non-spatial causal factor that nevertheless acted into space. He emphasized that it was a natural (as opposed to a metaphysical or mystical) factor that acted on physico-chemical processes. It was not a form of energy, and its action did not contradict the second law of thermodynamics or the law of conservation of energy. Then how did it work?
During the era of classical physics, when Driesch was writing, it was generally thought that all physical processes were fully deterministic, completely predictable in terms of energy, momentum, etc., at least in principle. But he considered that physical processes could not be fully determinate, because otherwise the non-energetic entelechy could not act upon them. He therefore concluded that, at least in living organisms, microphysical processes were not fully determined by physical causality, although, on average, physico-chemical changes obeyed statistical laws. He suggested that entelechy acted by affecting the detailed timing of microphysical processes, by suspending them and releasing them from suspension whenever required for its purposes:
This faculty of a temporary suspension of inorganic becoming is to be regarded as the most essential ontological characteristic of entelechy ... Entelechy, according to our view, is quite unable to remove any kind of ‘obstacle’ to happening ... for such a removal would require energy, and entelechy is non-energetic. We only admit that entelechy may set free into actuality what it has itself prevented from actuality, what it has suspended hitherto.
Although this bold proposal of a physical indeterminism within living organisms seemed to be completely unacceptable from the point of view of orthodox physics, it is much less outrageous in the light of quantum theory. Heisenberg put forward the uncertainty principle in 1927, and it soon became clear that positions and timings of microphysical events could be predicted only in terms of probabilities. By 1928, the physicist Sir Arthur Eddington was able to speculate that the mind influences the body by affecting the configuration of quantum events within the brain through a causal influence on the probability of their occurrence. “Unless it belles its name, probability can be modified in ways which ordinary physical entities would not admit of”. Comparable ideas were proposed by the neurophysiologist Sir John Eccles, who summarised his suggestion as follows:
The neurophysiological hypothesis is that the ‘will’ modifies the temporal activity of the neuronal network by exerting spatio-temporal ‘fields of influence’ that become effective through this unique detector function of the active cerebral cortex. It will be noted that the ‘will’ or ‘mind influence’ has itself some spatio-temporal patterned character in order to allow it this operative effectiveness.
A number of similar but more detailed proposals have been put forward by physicists and by parapsychologists.
However, even if entelechy could act by patterning probabilistic events, this hypothesis is unsatisfactory simply because it is vitalistic. Entelechy is essentially non-physical, by definition; even though it could, ex hypothesi, act on probabilistic events, this would still be an action of unlike on unlike. The physical world and non-physical entelechy could never be explained or understood in terms of each other.
This dualism, inherent in all vitalist theories, seems particularly arbitrary in the light of the discovery of the self-assembly of structures as complex as ribosomes and viruses, indicating a difference of degree, and not of kind, from crystallization. Although the self-organisation of living organisms as a whole is more complex than that of ribosomes or viruses, there is sufficient similarity to suggest that here again is a difference of degree. This, at any rate, is what both mechanists and organicists prefer to think.
Possibly a vitalist theory would have to be accepted if no other satisfactory explanation of the phenomena of life were conceivable. In the early part of the twentieth century, when vitalism seemed the only alternative to the mechanistic theory, it gained considerable support in spite of its essential dualism. But the development of the organismic theory over the last 100 years has provided another possibility which has effectively superseded vitalism by incorporating many of its aspects within a larger perspective.
Organicism
Organismic theories of morphogenesis developed under a variety of influences: some from philosophical systems, especially that of Alfred North Whitehead; some from physics, in particular from the field concept; others from Gestalt psychology, itself strongly influenced by the concept of physical fields; and some from the vitalism of Driesch. These theories deal with the same problems that Driesch claimed were insoluble in mechanistic terms - regulation, regeneration and reproduction - but whereas Driesch proposed the non-physical entelechy to account for the properties of wholeness and directiveness exhibited by developing organisms, organicists proposed morphogenetic (or embryonic, or developmental) fields.
This idea was put forward independently by Alexander Gurwitsch in Russia in 1907, who was much influenced by Driesch, and later by Hans Spemann in Germany in 1924 and Paul Weiss in Austria in 1926. All were leading developmental biologists, and Spemann was awarded the Nobel Prize in 1935. However, apart from stating that morphogenetic fields played an important role in the control of morphogenesis, none of them specified how these fields worked. The field terminology was soon taken up by other developmental biologists, but it remained ill-defined, although it served to suggest analogies between properties of living organisms and inorganic electro-magnetic systems. For example, if an iron magnet is cut into two parts, two whole magnets are produced, each with their own magnetic field, rather than two half magnets, one with a North pole and the other with a South pole. This resembles the way that a dragonfly egg can regulate, in such a way that half an egg can give rise to a small but complete dragonfly embryo. If two magnets are brought together in the right orientation, they form a single magnet with a unified magnetic field, just part of one plant can be grafted into another and form a unified whole, as in an apple tree, whose roots and shoots come from different plants.
The British biologist Conrad Hal Waddington suggested an extension of the idea of the morphogenetic field to take into account the temporal aspect of development. He called this new concept the chreode (from the Greek chre, it is necessary, and hodos, route or path) and illustrated it by means of a diagram of an “epigenetic landscape”.

Figure: Waddingtons’ model of an epigenetic landscape
In this model the path followed by the ball as it rolls downwards corresponds to the development of a particular part of an egg. As development proceeds, a branching series of alternative paths are represented by the valleys. These correspond to development of the different types of organ, tissue and cell. In the organism these are quite distinct; for example, the kidney and liver have definite structures. They do not grade into each other through a series of intermediate forms. Development is canalized towards definite end points. Genetic changes or environmental perturbations may push the course of development (represented by the pathway followed by the ball) away from the valley bottom up the neighbouring hillside, but unless it is pushed above the threshold into another valley, the process of development will find its way back. It will not return to the point from which it started, but to some later position on the canalized pathway of change and reach the same end-point as usual. This represents regulation.
The concept of the chreode is very similar to that of the morphogenetic field, but it makes explicit the dimension of time that is only implicit within the latter.
Both these concepts were taken further by the mathematician René Thom as part of a comprehensive attempt to create a mathematical theory embracing morphogenesis, behaviour and language. His main concern was to find an appropriate mathematical formalism for these problems, which had so far resisted mathematical treatment. The aim was to produce mathematical models that correspond as closely as possible to developmental processes. These models are topological, qualitative rather than quantitative, and do not depend on any scheme of causal explanation. As he put it in his book Mathematical Models of Morphogenesis (1983), “One essential feature of our use of local models is that it implies nothing about the ‘ultimate nature of reality’; even if this is ever revealed by analysis complicated beyond description, only a part of its manifestation, the so-called observables, are finally relevant to the macroscopic description of the system. The phase space of our dynamical model is defined using only these observables and without reference to any more or less chaotic underlying structures.”
Thom’s mathematical models were dynamic in the mathematical sense of the word. Dynamic systems evolve towards attractors, and Thom explicitly connected his models with Waddington’s idea of chreodes, with ends or goals towards which systems develop. Both Thom’s and Waddington’s models are teleological, like Driesch’s concept of entelechy. This is one fundamental way in which vitalism and organicism both differ from the mechanistic theory, which denies that there is any causal role of ends or goals, with pull from the future, and assumes that all causation involves only pushing from the part.
Organicism and vitalism have much in common. They include the idea that development is guided or canalized in space and time by top-down causal factors that include within themselves the end or goal of the developmental process and thereby provide a way of thinking about regulation and regeneration. The main difference is that whereas Driesch tried to say how the process he proposed might work, Waddington and Thom did not. The concept of the chreode was therefore less open to attack because it remained so vague. In fact, Waddington regarded the concepts of chreodes and morphogenetic fields as “essentially a descriptive convenience”. Like several other organicists, he was afraid of being accused of vitalism, the deadliest of heresies, and denied that he was suggesting the operation of anything other than known physical causes.
However, not all organicists made this denial; some left the question open. This explicitly non-committal attitude is illustrated by the following discussion of the morphogenetic field by the developmental biologist Brian Goodwin:
One aspect of the field is that electrical forces can affect it. Other developing and regenerating organisms have also been found to have interesting and significant electrical field patterns, but I would not wish to suggest that the morphogenetic field is essentially electrical. Chemical substances also affect polarity and other spatial aspects of developing organisms; and again I would not wish to draw the conclusion that the morphogenetic field is essentially chemical or biochemical in nature. My belief is that its investigation should proceed on the assumption that it could be any, or all or none of these things; but that, despite agnosticism regarding its material nature, it plays a primary role in the developmental process.
Michael Levin, who is one of the leading contemporary proponents of the morphogenetic field concept, also leaves open the nature of the “biofields”, although he often emphasises their electrical aspect.
The openness of this concept makes it the most promising starting point for a detailed organismic theory of morphogenesis. But clearly, if morphogenetic fields are assumed to be fully explicable in terms of known physical principles, they represent nothing but an ambiguous terminology superimposed upon a sophisticated version of the mechanistic theory. Only if they are assumed to play a causal role, at present unrecognised by physics, can a testable theory be developed.
Exploring the nature of morphogenetic fields takes on a new urgency in the light of modern evolutionary developmental biology. With the rise of molecular biology from the 1960s to the 1990s, the concept of morphogenetic fields was eclipsed in favour of genes. But as the limitations of the molecular approach became increasingly apparent, fields have re-emerged as a central concept for the understanding of development. The formation of entire structures such as wings or antennae in fruit flies can be “switched on” or “switched off” by mutations in homeobox genes. The morphogenetic field behaves as a whole, and the genetic switches are often referred to as “field selector genes”. Development is modular.
In an evolutionary context, morphogenetic fields take on an even more important explanatory role. As Scott Gilbert and his colleagues have argued:
Homologous developmental pathways… are seen in numerous embryonic processes, and they are seen occurring in discrete regions, the morphogenetic fields. These fields (which exemplify the modular nature of developing embryos) are proposed to mediate between genotype and phenotype. Just as the cell (and not its genome) functions as the unit of organic structure and function, so the morphogenetic field (and not the genes or the cells) is seen as a major unit of ontogeny [development] whose changes bring about changes in evolution.
So, what are these fields, and how do they work? Some biologists and mathematicians avoid proposing any new physical principles and follow the example of René Thom in regarding them as aspects Platonic Forms, timeless principles of order in a transcendent realm beyond space and time. In his recent work, Michael Levin has also adopted a Platonic approach. But the problem with this idea of eternal, transcendent Forms, Ideas or changeless mathematical formulae is that they do not evolve, being eternal. Yet all biology, and indeed the entire cosmos is evolutionary. What happens now depends on what has happened before. Evolution is creative and depends on its own history, like human societies and cultures, religions, science and technology.
In my hypothesis of formative causation, I propose that morphogenetic fields are not outside space and time in a Platonic realm, but inside space and time: they are immanent in nature, not transcendent. I suggest they are inherited from previous similar systems by process called morphic resonance, a transmission of form and pattern across space and time, from the past to the present, based on similarity. As a result of morphic resonance, each species has a kind of collective memory. The laws of nature are more like habits than a kind of mathematical Napoleonic Code, imposed on the universe at the instant of the Big Bang. This hypothesis is described in my books A New Science of Life (re-titled Morphic Resonance in the US) and The Presence of the Past and in my online course on morphic resonance.
I plan to write more about morphic resonance in a future essay and show how it suggests a new understanding of morphogenesis and of evolution, as well as providing a testable organismic approach not only to biology, but to physics chemistry, psychology and the social sciences.
This reads like a master work in making mincemeat out of "the other guys."
Now that he's reduced them to rubble, behold the truth: It's morphic resonance, stupid.
Lol
TBH, I kind wanted more about morphic resonance.
I am very happy you are publishing regularly on Substack, Dr. Sheldrake. I am really trying to dial in my understanding of the theory of morphic resonance.